
El difícil equilibrio entre la pérdida de agua y la fijación de carbono en plantas / Juan Pedro Ferrio

Juan Pedro Ferrio
Unidad de Recursos Forestales
![]() Centro de Investigación y Tecnología Agroalimentaria de Aragón (CITA)
Centro de Investigación y Tecnología Agroalimentaria de Aragón (CITA)
Fundación Agencia Aragonesa para la Investigación y el Desarrollo (ARAID)
jpferrio@cita-aragon.es
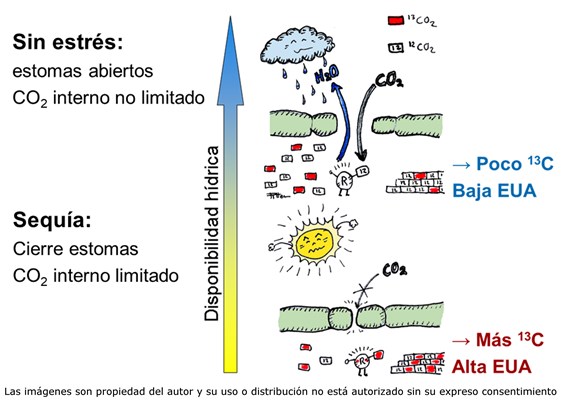
Desde la aparición de las primeras plantas terrestres, hace unos 500 millones de años, una de las principales “preocupaciones” de los vegetales ha sido la pérdida de agua. Las plantas superiores han desarrollado diversas estructuras que limitan la libre evaporación del agua (epidermis, cutícula, corteza). Así, las pérdidas de agua se concentran en las hojas y, más específicamente, a través de pequeñas aperturas en la epidermis, de diámetro regulable: los estomas. Mediante los estomas, las plantas pueden regular sus pérdidas de agua en función de las condiciones ambientales. Sin embargo, la limitación de las pérdidas de agua mediante el cierre de estomas tiene una contrapartida importante, ya que al reducir el intercambio con la atmósfera se restringe también el acceso del CO2 a los cloroplastos, y por tanto la fotosíntesis. Aunque existen mecanismos de concentración de CO2 (vías C4 y CAM), la mayor parte de las especies vegetales (plantas C3) carecen de estas adaptaciones, y se enfrentan diariamente a este dilema. Aquí es donde entra en juego un concepto fundamental en la ecofisiología vegetal: la eficiencia en el uso del agua, definida como la relación entre el carbono fijado y el agua transpirada (EUA). Aunque la regulación de los estomas permite modular la transpiración de las plantas, la tasa de pérdida de agua depende también de la demanda de agua del ambiente. Por esta razón, cuando comparamos distintas especies o genotipos, nos centramos en la eficiencia “intrínseca” en el uso del agua (EUAi), que es el cociente entre el carbono fijado y la conductancia estomática, y nos indica la facilidad con la que una planta deja escapar el agua.
La EUAi se puede determinar mediante medidas de intercambio de gases (flujo de CO2 y vapor de agua entre la planta y el aire) aunque esto proporciona un valor instantáneo, por lo que debe realizarse repetidas veces. Una alternativa muy extendida es el empleo de la composición isotópica de carbono (δ13C), ya que permite integrar, a partir de una muestra de material vegetal (hojas, semillas, madera), la EUAi a lo largo del período en que se formó el tejido. El principio de esta técnica es que las plantas fijan preferentemente el Carbono 12 (12C), frente al isótopo más pesado, el Carbono 13 (13C). Ambos isótopos son estables (no decaen con el tiempo, como el Carbono 14), pero su proporción se modifica por procesos físicos y químicos, lo que llamamos fraccionamiento. En el caso que nos ocupa, el 12CO2, más ligero y energético, difunde más fácilmente a través de los estomas que el 13CO2, pero también es procesado con mayor afinidad por la encima fijadora del carbono, RubisCO. El fraccionamiento (o discriminación) en contra del 13CO2 durante la fijación del CO2 por RubisCO es unas 6 veces mayor que el debido a la difusión, con lo que la importancia relativa de estos dos procesos modifica la composición isotópica de los azúcares fijados por la planta. Esto hace que, en especies con metabolismo C3, la δ13C sea un reflejo de la relación entre asimilación y conductancia estomática, y por tanto de la EUAi. Así, mediante el análisis de la δ13C es posible caracterizar la EUAi en un gran número de genotipos, tanto en programas de mejora como en la caracterización genética de especies forestales. Dado que la EUAi es un cociente entre dos variables, una mayor eficiencia puede asociarse a dos escenarios posibles, de consecuencias opuestas en términos de productividad.
En un primer escenario, el aumento de la EUAi se debe a una disminución de la conductancia estomática, como estrategia para limitar las pérdidas de agua. Esta limitación afecta negativamente a la fotosíntesis, por lo que la mayor eficiencia tendrá como contrapartida un descenso del crecimiento y la producción. Este es el caso, por ejemplo, de los cereales de invierno (trigo, cebada), donde los genotipos más productivos siguen una estrategia “despilfarradora” y son menos eficientes. Incluso en condiciones de sequía moderada, estos genotipos presentan rendimientos superiores a los de las variedades más “ahorradoras”. Únicamente en los ambientes más extremos, donde el rendimiento viene condicionado por la supervivencia, se han observado mejores rendimientos con la estrategia conservadora. No es de extrañar, por tanto, que en los programas de mejora de cereal se aplique de forma más o menos rutinaria la δ13C como criterio de selección negativo, identificando los genotipos con menor EUAi como aquellos con un mayor potencial productivo.
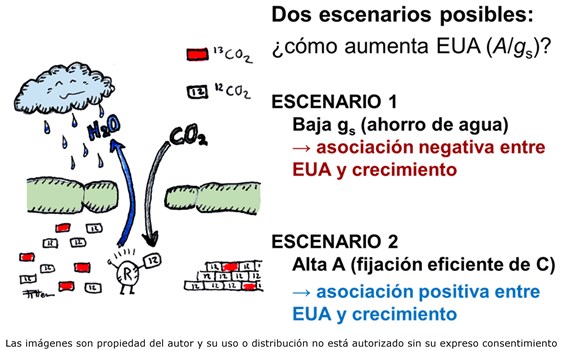
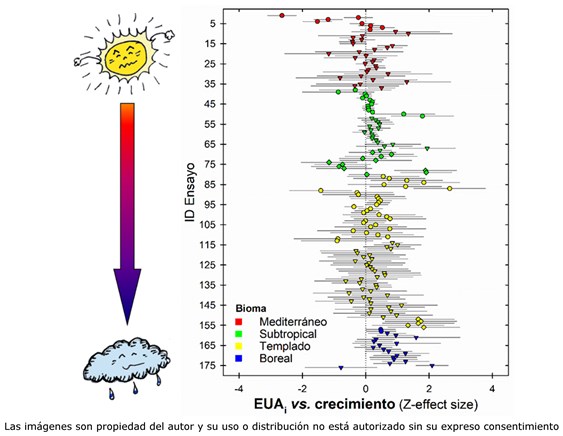
En un segundo escenario, la EUAi se incrementa gracias a una mayor capacidad fotosintética. En este caso, es de esperar una asociación positiva entre productividad y eficiencia hídrica. A diferencia de lo observado en cereales, ésta podría ser la estrategia dominante entre las especies forestales de interés comercial. En un meta-análisis a escala global, encontramos que la variabilidad genética en δ13C (y por ende la EUAi) se asocia positivamente con variables de crecimiento (altura, biomasa, diámetro). A diferencia de los cereales, originarios de zonas con alta irradiación solar y escasez periódica de agua, las especies forestales más productivas proceden en su mayoría de climas templados y boreales, donde la luz es tanto o más limitante que el agua. Bajo estas condiciones, la estrategia “ganadora” es aquella que permita optimizar la captación de luz. Por otro lado, en el contexto altamente competitivo del bosque, aumentar la fotosíntesis a nivel de hoja no es suficiente, y debe acompañarse de un crecimiento vigoroso, que permita competir por la luz. Esto podría haber reforzado la tendencia hacia una co-evolución positiva entre EUAi y crecimiento. No obstante, en algunas especies adaptadas a ambientes más secos (pino carrasco, nogal, eucalipto) se ha observado una asociación negativa entre δ13C y crecimiento, probablemente como respuesta a una presión selectiva en favor de estrategias más conservadoras bajo condiciones de estrés.
Aunque los dos escenarios descritos pueden parecer contradictorios, hay razones para ser optimistas, ya que existe margen para desarrollar cultivos más sostenibles, aumentando la eficiencia hídrica sin perder productividad. Entre los caracteres fisiológicos más prometedores en este sentido se encuentra la limitación de la fotosíntesis asociada a la difusión interna del CO2. Una vez atraviesa los estomas, el camino del CO2 hasta llegar al cloroplasto es relativamente independiente del flujo del agua. Por tanto, existen mecanismos para aumentar la difusión interna del CO2 (y con ello la fotosíntesis) sin que ello implique incrementar las pérdidas del agua. Una vez más, la mejora puede beneficiarse del conocimiento básico generado por la fisiología vegetal, que ha mostrado la enorme diversidad existente entre las especies silvestres.